植物生长素(IAA)在植物生长发育过程中起着重要的作用。其化学本质是吲哚乙酸。主要作用是使植物细胞壁松弛,从而使细胞生长伸长,在许多植物中还能增加RNA和蛋白质的合成。
目前BARLEY B RECOMBINANT/BASIC PENTACYSTEINE (BBR/BPC) 转录因子家族在拟南芥和水稻中已被证实能促进诱导植物矮化(Sazuka et al. 2009; Li et al. 2008Monfared et al. 2011; Simonini et al. 2012; Petrella et al. 2020),然而在苹果中,BPC转录因子影响植物结构的确切机制尚不清楚。
2023年11月29日,西北农林科技大学园艺学院的李明军教授课题组的研究成果,发表在The Plant cell期刊上(IF=11.6),文章题目是“The transcription factor MdBPC2 alters apple growth and promotes dwarfing by regulating auxin biosynthesis”。该研究使用DNA亲和测序(DAP-seq)技术和转录组测序(RNA-seq)技术鉴定及揭示了BBR/BPC家族的MdBPC2转录因子能够调控苹果生长素生物合成,从而协调苹果的生长和植株结构的机制。

本研究发现MdBPC2转录因子在矮秆砧木中的表达高于标准砧木。MdBPC2与PcG蛋白MdLHP1a/b相互作用,负向调节生长素的生物合成。在过表达MdBPC2的转基因苹果植株中,抑制MdYUC2a和MdYUC6b基因导致生长素积累减少,植株生长受到抑制,树形结构发生改变。外源生长素的补充恢复了这些转基因苹果植株的高度、叶片形态和根系发育。在WT植株中,抗生长素对氯苯氧异丁酸(PCIB)处理抑制了生长素的生物合成,导致与MdBPC2过表达转基因苹果植株相似的表型。最终揭示了MdBPC2如何通过H3K27me3修饰调控生长素的生物合成以协调苹果的生长和植株结构的机制。
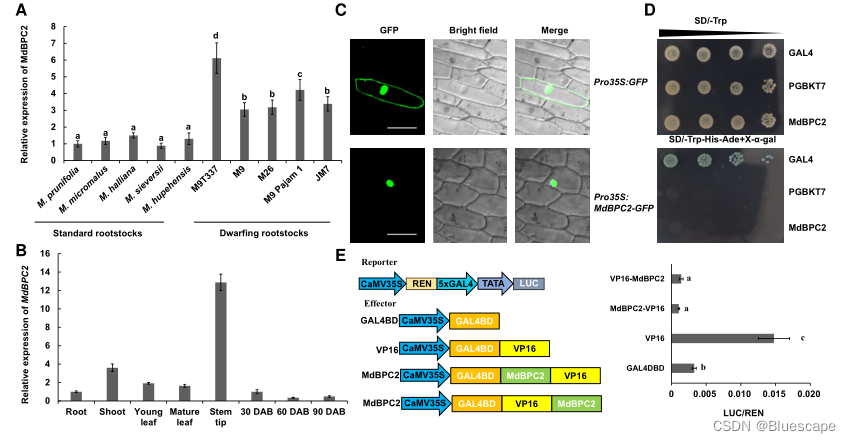
1.MdBPC2在矮秆砧木中的表达及特性研究。
作者首先在苹果中鉴定出6个MdBPC候选基因,并在系统发育树中分为2类,MdBPC1与MdBPC2序列高度相似,属于一类。MdBPC3、MdBPC4、MdBPC5、MdBPC6属于二类。为确定MdBPC是否具有调控植物生长的作用,比较了6个MdBPC基因在5个矮化砧木和5个标准砧木中的表达谱。I类BPCs的表达水平,尤其是MdBPC2在矮秆砧木中的表达水平显著高于标准砧木。因此,最终选择MdBPC2作为进一步研究的候选基因。
利用定量PCR (qPCR)研究了MdBPC2基因在不同组织中的表达谱。结果表明,MdBPC2在整个植物中广泛表达,其中茎尖表达量较高。为确定MdBPC2的亚细胞定位,瞬间表达了MdBPC2-GFP融合蛋白,表明了MdBPC2在细胞核中起作用。通过酵母杂交实验,表明MdBPC2没有转录激活活性,提示MdBPC2可能是一种转录抑制因子。利用荧光素酶(luciferase, LUC)报告系统分析了其在植物中的转录活性,结果表明MdBPC2具有转录抑制活性。

2.过表达MdBPC2和RNAi转基因苹果株系的鉴定及其对植株生长的影响。
作者构建了过表达和RNA干扰(RNAi)载体,并将其转化为苹果品种GL3。PCR分析得到3个过表达系(MdBPC2-OE1/3/4)和3个RNAi系(MdBPC2-RNAi3/5/6)。反转录定量PCR (RT-qPCR)分析显示,与WT植株相比,转基因MdBPC2-oe1/3/4植株的MdBPC2转录本约高15倍,转基因MdBPC2-RNAi3 /5/6植株的MdBPC2转录本约低60%。之后还测量了MdBPC2转基因苹果植株中其他5个MdBPC基因的表达水平。在MdBPC2沉默系中,MdBPC1的转录水平也受到抑制,可能是由于序列相似性较高。在MdBPC2过表达系中,植株生长明显受到抑制。MdBPC2过表达显著降低了植株的平均株高,仅为WT植株的29%。叶片形态也发生了变化,长宽比减小,形状更圆。此外,过表达植株根系发育不发达,根长缩短,侧根数量减少。然而,在MDBPC2沉默系中,没有明显的表型变化,这可能是由于BPC家族成员之间的功能冗余。
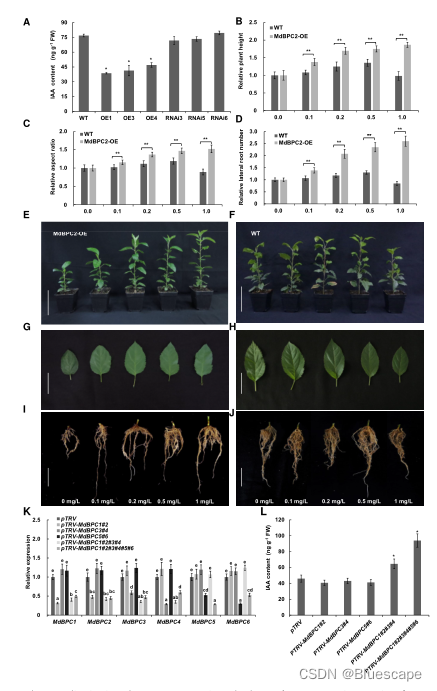
3.MdBPC2改变生长素含量
植物激素调节植物的生长发育,内源激素含量或平衡的改变可能导致植物矮化。因此,作者在转基因过表达系和RNA沉默系中测量了生长素(IAA)、细胞分裂素(CTK)、GA和油菜素内酯(BR)等激素含量。与WT相比,CTK、GA和BR的含量没有变化,而MdBPC2过表达系的IAA含量显著降低。为进一步确定MdBPC2-OE表型是否由IAA缺乏引起,作者用外源IAA处理了MdBPC2-OE转基因植株,外源施用IAA能以剂量依赖的方式恢复MdBPC2-OE植株的株高、叶片发育和根系生长。重要的是,IAA促进了MdBPC2-OE植株的生长,而WT植株的表型不受IAA处理的影响。这表明MdBPC2-OE植株对IAA浓度高度敏感。之后增加抑制生长素作用的PCIB浓度处理WT植株,植株高度降低,叶片形态改变,侧根数量显著减少,且呈剂量依赖性。结果表明,MdBPC2过表达导致内源生长素含量降低,从而抑制转基因植株的生长。
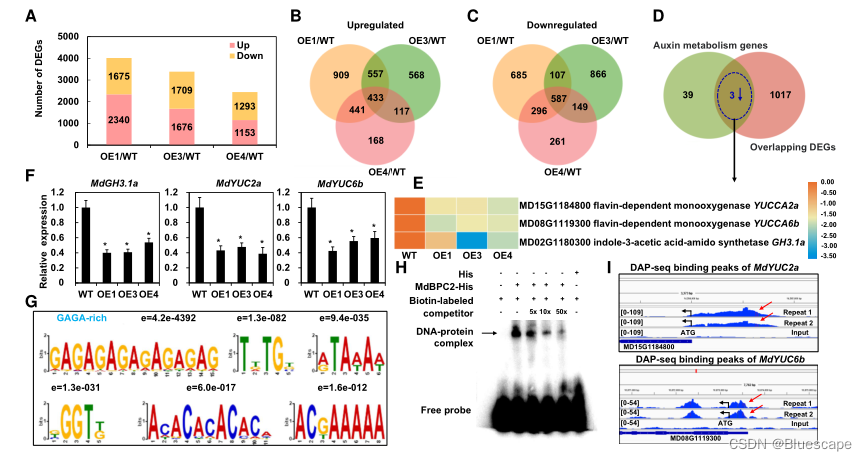
4.MdBPC2直接靶点的鉴定
为确定MdBPC2过表达如何改变内源IAA积累,作者使用WT和MdBPC2-OE1/3/4植株的茎尖进行了RNA测序(RNA-seq)。鉴定出433个上调基因和587个下调基因(与WT相比)。已知参与生长素合成和分解代谢的基因与MdBPC2-OE转基因品系的1020个基因进行比较,共发现3个重叠基因,分别是MdYUC2a和MdYUC6b以及MdGH3.1a,并且在3个转基因株系中表达下调。为了验证RNA-seq表达谱数据,通过qRT-PCR分析了这3个基因,发现表达趋势与转录组数据一致,这也说明了RNA-seq技术的准确性。
DNA亲和纯化测序(DAP-seq)是一种通过分析基因组DNA与体外表达的转录因子的结合来筛选转录因子结合位点的高通量方法。与染色质免疫沉淀(ChIP-seq)相比,具有具有不受抗体和物种限制的优点,是一种高通量方法。因此,作者使用DAP-seq来揭示MdBPC2的全基因组结合位点,分离所有MdBPC2结合DNA进行高通量测序分析。“GAGAGAGAGAGAG”这个富含GAGA的核苷酸序列比拟南芥BPC1结合位点更为保守。另外5个MdBPC2的潜在结合位点也高度富集。
电泳迁移率转移试验(EMSA)验证DAP-seq结果的可靠性。当使用标记的富含GAGA的DNA探针时,可以检测到特异性DNA-MdBPC2蛋白复合物,当添加缺乏生物素标记的竞争性探针时,这种信号传导减弱。这种特异性竞争组合表明MdBPC2可以特异性识别富含GAGA的序列。结果表明,MdBPC2在转录水平上通过抑制MdYUC2a和MdYUC6b抑制生长素的生物合成。
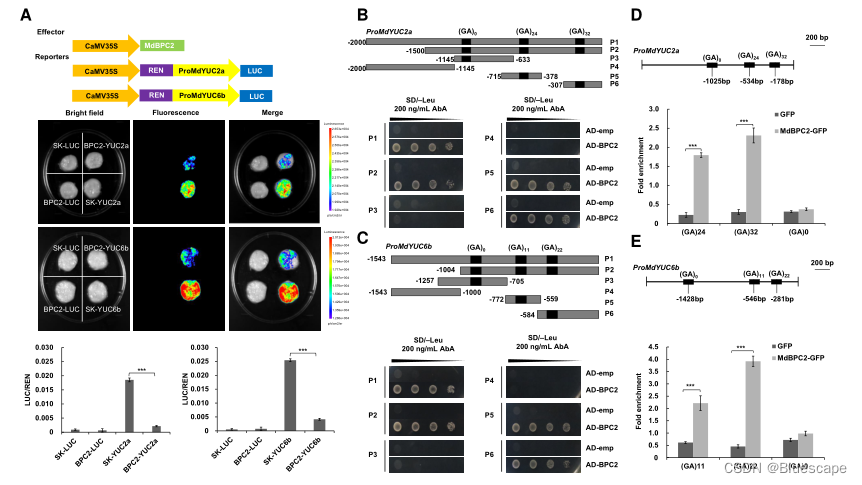
5.MdBPC2抑制MdYUC2a和MdYUC6b的转录
为确定MdBPC2是否直接影响MdYUC2a和MdYUC6b的转录激活,作者在苹果愈伤组织中瞬时表达系统中检测了ProMdYUC2a:LUC和ProMdYUC6b:LUC构建体的生物发光信号,结果表明,MdYUC2a和MdYUC6b启动子区域中所有富GA元件都可以与MdBPC2蛋白结合。
ChIP-qPCR结果表明MdBPC2可以通过GA-富元件与MdYUC2a和MdYUC6b启动子结合。这些结果表明,生长素生物合成的关键基因MdYUC2a和MdYUC6b是MdBPC2的直接转录靶点,其转录活性受到MdBPC2的抑制。
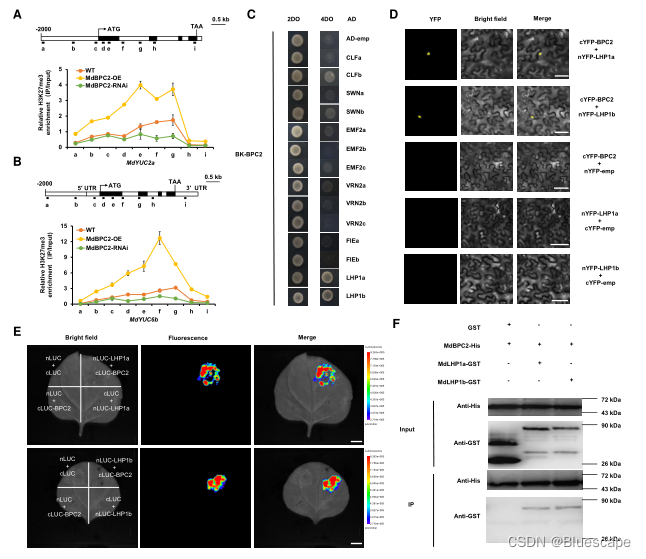
6.MdBPC2介导抑制组蛋白三甲基化在H3K27位点的富集
翻译后组蛋白修饰与基因转录的激活或抑制有关。组蛋白3赖氨酸27三甲基化(H3K27me3)的全基因组研究发现,表观遗传机制可以调节生长素相关基因。为检测H3K27me3抑制标记是否与MdBPC2对MdYUC2a和MdYUC6b的抑制有关,通过ChIP检测MdBPC2过表达后H3K27me3抑制标记在MdYUC2a和MdYUC6b位点的潜在差异,结果表明MdBPC2在抑制MdYUC2a和MdYUC6b表达的表观遗传修饰中起重要作用。
7.BPC2蛋白与PcG亚基LHP1a/b物理相互作用
在植物中,抑制表观遗传修饰H3K27me3主要由PcG蛋白识别和催化。作者使用酵母双杂交(Y2H)实验来测试MdBPC2与抑制性表观遗传修饰相关蛋白的相互作用,包括MEDEDA (MEA)、CURLY (CLF)、SWINGER (SWN)、胚胎花2 (EMF2)、受精独立胚乳(FIE)和LIKE异染色质蛋白1 (LHP1)。研究结果表明,MdBPC2与MdLHP1a/b相互作用。通过双分子荧光互补(BiFC)进一步评估。通过共表达融合蛋白nYFP-LHP1a/b和cYFP-BPC2,能在烟草表皮细胞细胞核中重建YFP的活性,证明MdLHP1a/b与MdBPC2相互作用。这些结果表明,MdBPC2可以通过MdLHP1a/b募集PcG复合物到靶基因。
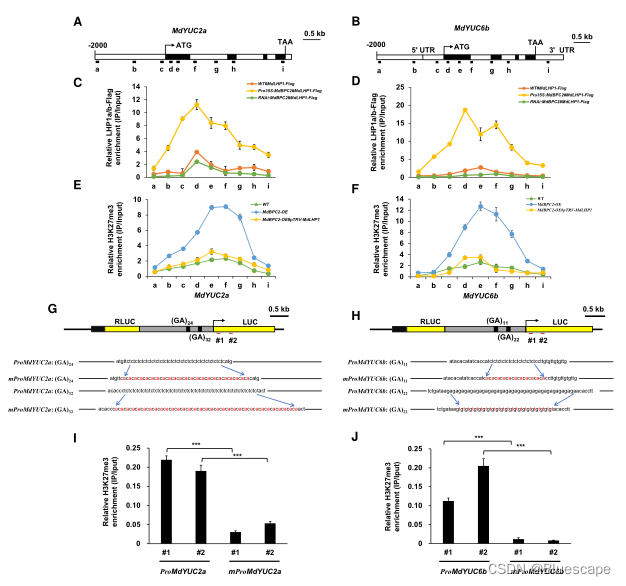
8.LHP1参与MdBPC2介导的YUC2a/6b表达抑制
由于MdBPC2可以特异性结合GAGA序列,并且MdBPC2和MdLHP1之间存在相互作用,作者假设MdLHP1在MdBPC2诱导的MdYUC2a和MdYUC6b表达抑制中起作用。为验证这一假设,首先使用ChIP-qPCR检测了MdYUC2a和MdYUC6b位点富集的MdLHP1水平,结果显示MdYUC2a和MdYUC6b位点H3K27me3水平的变化是由MdLHP1引起的。
为验证MdLHP1在MdYUC2a和MdYUC6b启动子上的富集是否需要MdBPC2的结合,制作了4个转基因苹果愈伤组织系,含有突变的富GA元素的转基因品系与含有完整的富GA元素的转基因品系相比,H3K27me3水平显著降低。这些结果表明,靶启动子内的MdBPC2结合位点对于MdLHP1介导的H3K27me3修饰是必需的。
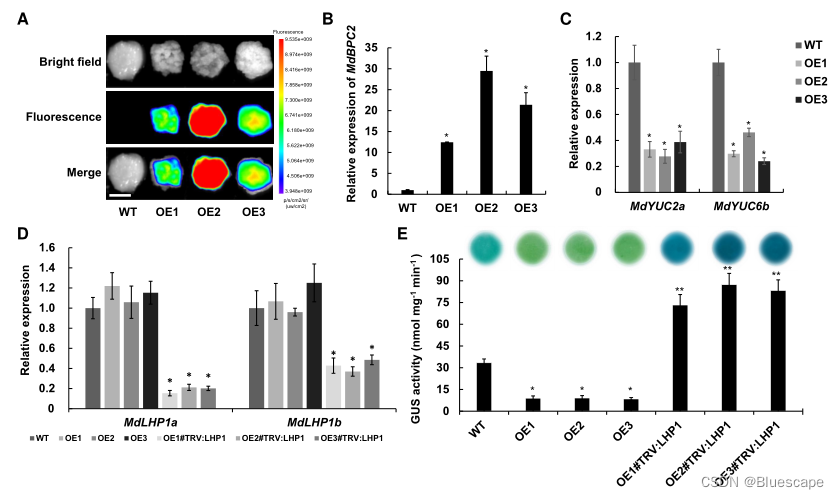
9.MdLHP1调控生长素生物合成基因
为证实MdLHP1参与了MdBPC2对生长素水平的调节在苹果愈伤组织中过表达MdBPC2,获得了3个转基因系MdBPC2-GFP-OE1/2/3。这些转基因系的愈伤组织中MdYUC2a和MdYUC6b的表达水平显著降低,与转基因植物中的观察结果一致。
DR5是一种合成的生长素反应元件,含有许多ARF转录因子的结合位点。因此,在DR5启动子控制下表达的报告基因活性可以反映植物内源生长素含量。利用合成的ProDR5:GUS构建体分析了转基因苹果愈伤组织MdBPC2-GFP-OE1/2/3的生长素含量。如图E所示,与WT相比,MdBPC2-GFP-OE1/2/3的GUS染色明显降低。当沉默MdLHP1时,GUS活性恢复并且高于WT。综上所述,这些数据表明MdLHP1参与了MdBPC2在生长发育过程中对生长素生物合成的调节。
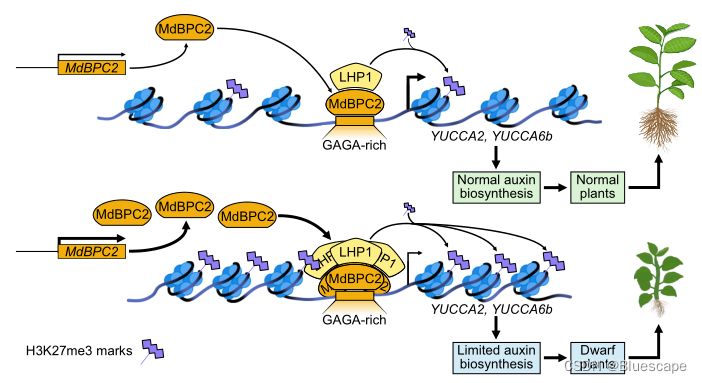
小结
株高是农作物和园艺作物最重要的农艺性状之一。它限制了植物各器官的空间排列,与产量和品质密切相关。植物高度受环境、植物激素代谢和信号通路的相互作用控制,转录因子在这些过程中发挥重要作用。虽然转录因子在植物高度调节中的重要性早已被提出,但它们如何调节植物矮化的潜在分子机制却知之甚少。在本研究中,作者发现MdBPC2调控MdYUC2a和MdYUC6b的转录,抑制生长素的生物合成,从而调节植物结构。揭示了多年生木本植物通过BPC转录因子调控生长素合成的新机制,为优化植物结构和提高产量提供了新的思路。


和整合ajax异常处理)
数字经济-215个工具变量汇总(更新至2024年))







)



)


。Javaee项目,ssm vue前后端分离项项目。)
