关于酵母双杂交服务的常见问题
问题1:酵母双杂交的筛选流程?
研究者将特定基因作为钓饵,在一个精心挑选的cDNA文库中进行筛选,目的是找到与该钓饵蛋白发生相互作用的蛋白质。通过这种筛选,可以从阳性反应的酵母菌株中提取出含有AD-LIBRARY质粒的样本。接下来,对这些质粒中的cDNA片段进行测序分析,并将其编码的序列与GENEBANK数据库中的序列进行对比,从而探究其与已知基因在生物学功能上的潜在关联。
问题2:酵母双杂交自激活原理
一般BD可单独与GAL4上游活化序列UAS结合,但不能引起转录。如果将一段具有转录激活活性的转录因子构建到BD载体上,若其表达产生的融合BD单独与UAS结合后能引起下游报告基因的转录,则说明bait protein具有自激活现象。
BD-bait单转化菌株要经过SD-Trp/-His/-Ade验证其是否存在自激活;再与AD空载体杂交后在SD-Trp/ -Leu/-His/ -Ade上进一步验证其与DNA激活domain是否存在直接互作。
AD-prey单转化菌株要经过SD-Leu/-His/-Ade验证其是否存在自激活,再与BD空载体杂交后在SD-Trp/ -Leu/-His/ -Ade上进一步验证其与DNA结合domain是否存在直接互作。
问题3:如何解决酵母双杂交实验中的自激活问题?
自激活是一个常见问题。可以通过更换酵母菌株、调整培养条件或添加竞争抑制剂来解决自激活问题。若诱饵蛋白具有自激活活性,能够单独激活报告基因则:(1)该蛋白是一个转录因子,具有转录激活域,需要去掉转录激活结构域。(2)该蛋白不是转录因子但仍具有较强的转录激活活性,需要将诱饵蛋白具有激活活性的区域去除,进行后续操作,但这样操作有可能影响蛋白间的互作。
问题4:获得的阳性克隆,PCR检测有多条片段
可能原因:一个酵母细胞可以包含多个捕获蛋白。解决办法:选取单克隆划板2-3次,进行蓝白斑筛选,直到没有分离,找到阳性克隆,用于后续实验。
问题5:AD和BD先转后转有区别吗?
有区别。如果先转AD,则需要涂-Leu平板;如果先转BD,则需要涂-Trp平板
问题6:在筛选过程中,如何确保筛选到的阳性克隆是真阳性,而非假阳性?如何降低酵母双杂交系统中出现的假阳性?
通过多次验证和不同条件下的重复实验,以及利用其他方法如共沉淀实验来验证阳性克隆的真实性。或者可以选择多个筛选标记进行更为严格的筛选,降低假阳性;或者通过适当提高3-AT的浓度(一般 10 mM浓度的3-AT平板会有少量生长,20 mM浓度的3-AT不能或极少生长。如果在80 mM浓度的3-AT平板上生长酵母,说明自激活活性太高,不能用于双杂交筛选。),降低假阳性。
问题7:酵母菌落长时间培养后变红
这是由于酵母菌株在低含量腺嘌呤培养基上生长时嘌呤前体积累造成的,一般不影响蛋白质互作实验。一般通过往培养基中补加硫酸腺嘌呤(60mg/L)来改善这一情况。
问题8:酵母细胞生长较慢
可能原因:酵母表达的诱饵蛋白对细胞有毒害作用,影响酵母生长。解决办法:可通过选择低敏感性的酵母菌株;或选用低拷贝数的表达载体。
问题9:在酵母双杂交实验中,如何解决低转化效率的问题?
(1)优化转化条件、使用新鲜的酵母菌株、载体以及提高DNA质量可以帮助提高转化效率。(2)在杂交中,预转化的诱饵细胞的数量可能不够。当对诱饵菌株进行液体培养过夜时,应挑选大的、新鲜的克隆进行培养,经过离心和重悬后,再使用血球计对细胞进行计数,提高杂交效率。
问题10:如何确定是选择构建核体系文库还是膜体系文库?
如果诱饵基因是定位在膜上的蛋白则选择膜体系文库,如果诱饵基因是定位于核则选择核体系文库,如果诱饵基因有跨膜区也可以考虑把跨膜区切除以后用核体系文库筛库,但是这样操作有一定风险。
问题11:为什么选择酵母作为双杂交系统的报道株?且选择合适的酵母菌株很重要?
酵母作为酵母双杂交系统的报道株,它具有诸多优点:易于转化、回收扩增质粒,具有可直接进行选择的标记基因和特征性报道基因;酵母内源性蛋白不易同来源于哺乳动物的蛋白结合。合适的酵母菌株能确保实验的准确性和重复性。选择具有所需表型特性和遗传背景的酵母菌株是非常重要的。
问题12:在酵母双杂交实验中,如何解决交叉反应问题,提高筛选的特异性和敏感性?
优化筛选条件,例如调整培养温度、时间和基质浓度,以及使用高特异性的抗体和优质的试剂可以提高筛选的特异性和敏感性。及通过其他方法如共免疫沉淀验证蛋白质相互作用的特异性,可以减少非特异性的蛋白质相互作用。
问题13:在筛选过程中,如何解决背景噪音问题?
调整培养条件、更换酵母菌株或优化实验设计可能会降低背景噪音,提高实验的准确性。
问题14:如何解决酵母双杂交实验中的蛋白质表达水平低的问题?
优化表达载体、更换强表达的启动子或者选择高效表达的酵母菌株,以及调整培养条件可能有助于提高蛋白质表达水平。
问题15:如果在筛选过程中遇到了高度相似的阳性克隆,如何区分它们的差异?
通过序列分析、功能验证和其他生物信息学方法来区分和解析高度相似阳性克隆的差异。
问题16:如何提高酵母双杂交实验的高通量筛选效率?
使用自动化的实验设备和优化筛选流程,以及使用高效的数据处理和分析方法可以提高高通量筛选效率。
问题17:在酵母双杂交实验中,如何解决蛋白质溶解度低的问题?
优化表达条件、使用溶解度增强型载体或添加溶解度增强剂,可能有助于改善蛋白质的溶解度。
问题18:在酵母双杂交实验中,如何提高蛋白质的折叠和功能活性?
优化表达和纯化条件,以及使用专门的折叠辅助剂和折叠缓冲液,可以提高蛋白质的折叠和功能活性。
问题19:如何解决酵母双杂交实验中的蛋白质降解问题?
使用蛋白质稳定性增强型的表达系统和载体,以及优化纯化条件和储存条件,可能有助于减少蛋白质降解。
问题20:酵母杂交技术有哪些优势?
(1)体内的互作验证,省去蛋白表达、纯化等步骤;(2)细胞内验证,在一定程度上反应细胞内的真实情况;(3)可以检测微弱的蛋白互作;(4)可对不同组织、器官、细胞、分化阶段材料进行文库构建和筛选。
卡梅德生物(KMD Bioscience)(https://www.kmdbioscience.cn/) 可提供酵母单/双杂交文库构建、酵母单/双杂交验证服务,卡梅德生物(KMD Bioscience)多年来致力于酵母双杂交系统(Yeast two-hybrid system,Y2H)的研究。我们的酵母杂交系统是由基于SMART法构建的文库、酵母菌株、严谨的报告基因、高表达载体组成的。这项技术可用于:研究蛋白的相互作用;确认蛋白的交互作用;定义蛋白的交互域。
案例
核体系酵母双杂案例:
核因子Y(NF-Y)是一种异源三聚体转录因子,由三个亚基组成,NF-YA、NF-YB和NF-YC。NF- Ys在胚乳发育过程中的基因功能尚不清楚。在这项研究中,鉴定了八个水稻NF-Y编码基因,即OsNF-YA8、OsNF-YB1、9和OsNF-YC8、9、10、11、12,这些OsNF-Y的紧密同源物仅存在于单子叶植物物种中,并且也优先在胚乳中表达,这表明它们在胚乳发育的调节中发挥作用。
为了验证这一想法,进行了酵母双杂交分析,以检测不同组胚乳优先OsNF- Ys之间的蛋白质相互作用。同源性建模分析显示,尽管序列可变,但蛋白质结构没有变化。NF-YB,包括OsNF-YB1、OsNF-YB9和OsNF-YB7,能够与任何胚乳优先的NF-YC家族成员相互作用。然而,OsNF-YA8只能与OsNF-YB9相互作用,尽管 OsNF-YB7 和 OsNF-YB9 高度相似,但 OsNF-YA8 在酵母中并未显示出与 OsNF-YB7 的相互作用。
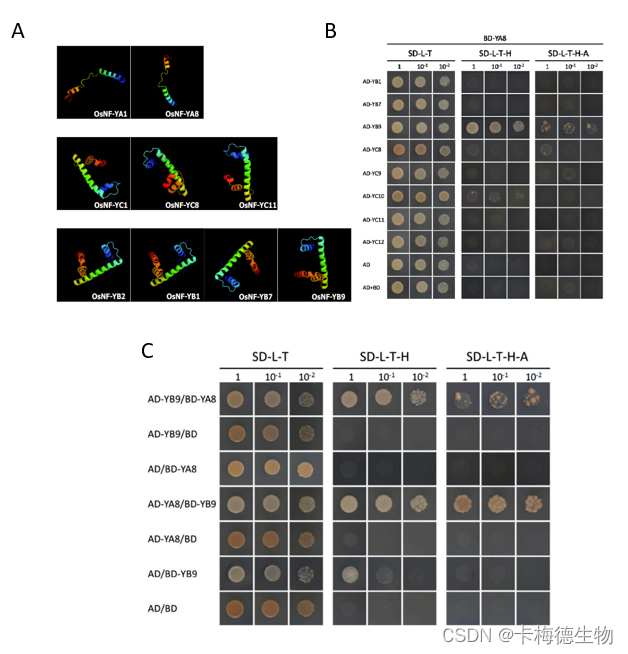
Fig.1 (A)Predicted structures of the NF‐Ys’ conserved domains. (B)Yeast-two-hybrid assays show interactions between OsNF-YA8 and OsNF-YB9. AD and BD indicate the activation domain and the DNA-binding domain of GAL4, respectively; AD-YA8 indicates the AD::OsNF-YA8 fusion, AD-YB9 indicates AD::OsNF-YB9, BD-YA8 indicates BD::OsNF-YA8, and BD-YB9 indicates BD::OsNF-YB9. Serial dilutions of the yeast cells expressing the indicated proteins were plated on non-selective (SD–L–T) or selective (SD–L–T–H and SD–L–T–H–A) medium.(C)OsNF‐YA8 interacts with OsNF‐YB9, but not with other endosperm‐preferential OsNF‐Ys.【A group of nuclear factor Y transcription factors are subfunctionalized during endosperm development in monocots.DOI:doi:10.1093/jxb/ery087】
核体系酵母双杂案例:
SKIP是许多真核生物中的转录辅因子。它可以调节水稻和拟南芥的植物抗逆性。但大豆SKIP蛋白的同源物至今尚未报道。在本研究中,大豆的GAMYB结合蛋白基因(GmGBP1)编码SKIP蛋白同源物在非生物胁迫和不同日照时间下的生长被分析。文献首先对GmGBP1自激活的检测。验证了GmGBP1在C端具有转录活性。
在大麦中,已经证实HvGAMYB可以通过酵母双杂交系统与SKIP结构域相互作用。因此,进行了酵母双杂交试验来研究GmGBP1和GmGAMYB1之间的相互作用。GmGAMYB1包含一个典型的R2/R3-MYB DNA结合结构域和三个保守区(BOX1、BOX2和BOX3)。由于GmGBP1在C末端结构域中显示出转录活性,因此选择pBD-GmGBP1a和pBD-Gm GBP1b作为诱饵载体,以验证与GmGAMYB1的互作。此外,猎物通过R2R3的结构域区域分为两部分(1–141,142-538),分别为pAD-Gm GAMYB1a和pAD-Gm-GAMYB1b。pBDGmGBP1a和猎物质粒的共转化不能在SD/TLH上生长,表明GmGBP1a与GmGAMYB1没有相互作用。此外,具有pBD-GmGBP1b和pAD-GmGAMYB1b的酵母细胞没有表现相互作用的能力。相反,由于在SD/-TLH上的生长,GmGBP1b通过与GmGAMYB1a的结构域结合而与GmGAMYB1相互作用,表现出阳性显色反应(X-Gal过滤分析中的蓝色,图3D)。
GmGBP1与酵母细胞中的GmGAMYB1相互作用:

Fig.1 (A)Detection for self-transactivation of GmGBP1. (B) The growth of Yeast YRG-2 cells cotransformated by pBD-GmGBP1a with pAD-GmGAMYB1, pADGmGAMYB1a (amino acids 1–141) or pAD-GmGAMYB1b (amino acids 142–538) on synthetic minimal (SD) medium lack of Trp and Leu (SD/-TL) or Trp, Leu and His (SD/-TLH). (C) The growth of Yeast YRG-2 cells cotransformated by pBD-GmGBP1b with pAD-GmGAMYB1, pAD-GmGAMYB1a or pAD-GmGAMYB1b on SD/-TL and SD/-TLH, and the X-Gal assay for the transformations.【GmGBP1, a homolog of human ski interacting protein in soybean, regulates flowering and stress tolerance in Arabidopsis.DOI:10.1186/1471-2229-13-21】
这篇文章可供科研爱好者参考。它不能代替需要更详细和专业信息的专业知识或实践实验程序。如果有任何内容侵权,请联系作者立即删除有争议的材料。



)







)

)
)

)


